

公募研究
B01: 暗所生育が可能なシアノバクテリアを活用したクロロフィル新機能と光化学系個別再構成
代表者

藤田 祐一(名古屋大学 大学院生命農学研究科 教授) ![]()
研究協力者
 山本 治樹(名古屋大学 大学院生命農学研究科 助教)
山本 治樹(名古屋大学 大学院生命農学研究科 助教)概要
シアノバクテリアは光合成の進化と仕組みを理解する上で重要な生物ですが、これまで多くの研究で用いられたシアノバクテリアは光栄養を生育の必須要件とするため、クロロフィルを完全に失うといった極端な変異株が単離できず、光合成をゼロの状態から改めて再構築させるような実験系を組むことができませんでした。シアノバクテリアLeptolyngbya boryanaは完全暗所でも糖を利用して従属栄養的に生育でき、光合成にまったく依存しない生育が可能です。本研究では、L. boryanaの高い従属栄養能を活用し、クロロフィルの新規機能、クロロフィルほぼゼロベースからの個別光化学系in vivo再構築そしてチラコイド膜形成に至る過程を統合的に明らかにし、酸素発生型光合成の進化に迫ります。
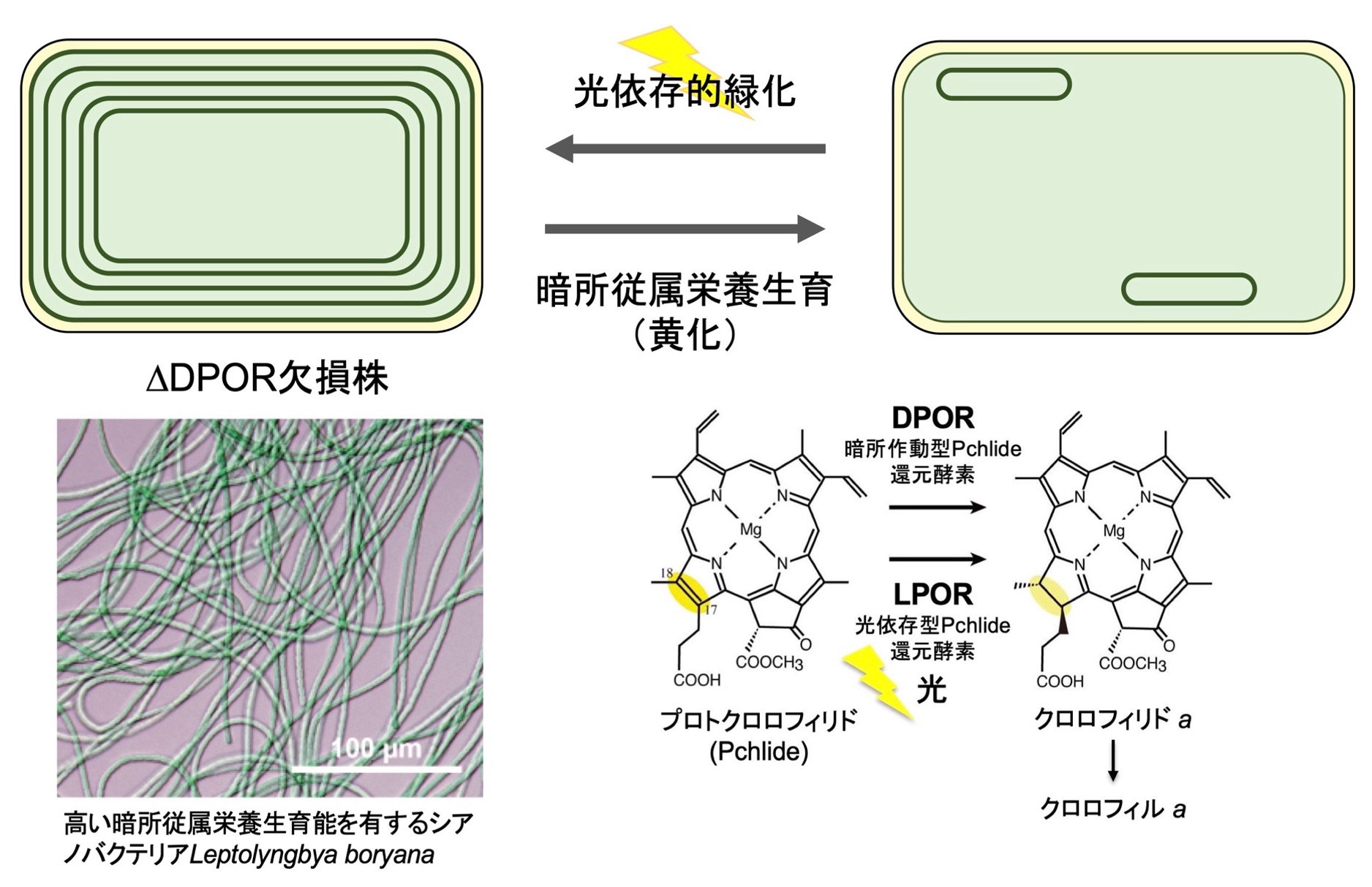
研究キーワード:シアノバクテリア、暗所従属栄養、クロロフィル生合成、チラコイド膜、光依存的緑化、黄化
B01: シトクロムb6f複合体のpH感受性最適化の分子進化
代表者

鹿内 利治(京都大学 理学研究科 教授)![]()
研究協力者

竹中 瑞樹(京都大学 理学研究科 准教授)![]()
概要
シトクロムb6f複合体は、光化学系Ⅱから光化学系Ⅰへの電子伝達を触媒し、Qサイクルと呼ばれる機能で、プロトン勾配の形成に機能します。電子伝達によって生じるチラコイド膜ルーメンの酸性化は、シトクロムb6f複合体の活性を抑制することで、下流の光化学系Ⅰに過剰な電子が流れることを防いでいます。このことは、自然光の様に強度が変動する光環境の下で、光化学系Ⅰを光傷害から防ぐのに必須です。我々は、pH感受に関わると考えられる複合体のRieskeサブユニットに一連の変異を導入し、複合体での電子伝達ブレーキの強さを最適化する分子進化について知見を得ることを目指します。
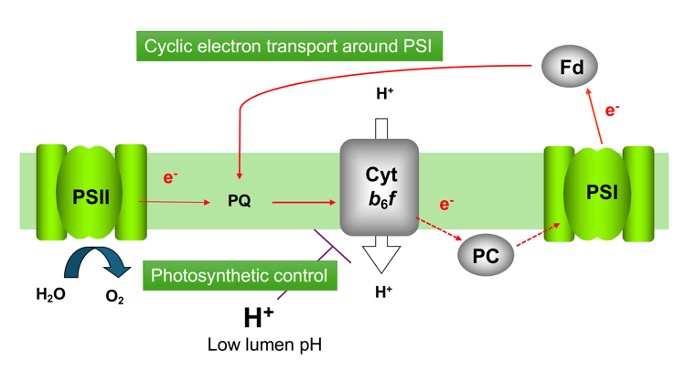
研究キーワード:シトクロムb6f複合体、光合成、光化学系Ⅰ、サイクリック電子伝達
B01: 構造から探る酸素非発生型光合成におけるNPQの生理機能
代表者

浅井 智広(中央大学 理工学部 准教授)![]()
研究協力者

小澄 大輔(熊本大学 産業ナノマテリアル研究所 准教授)
研究協力者

稲垣 知実(立命館大学 大学院生命科学研究科 博士後期課程)
概要
絶対嫌気性の光合成細菌である緑色硫黄細菌では,光合成反応中心複合体内部でカロテノイド配糖体が非光化学的消光(NPQ)を駆動していることがわかっています。これは,活性酸素種(ROS)の生成が関与しない,「嫌気的な光阻害現象」が存在することを示しています。私たちは,緑色硫黄細菌の光合成反応中心複合体で起こるNPQの反応メカニズムを原子レベルの詳細さで解析し,無酸素環境下でのカロテノイドやNPQの存在意義を明らかにします。
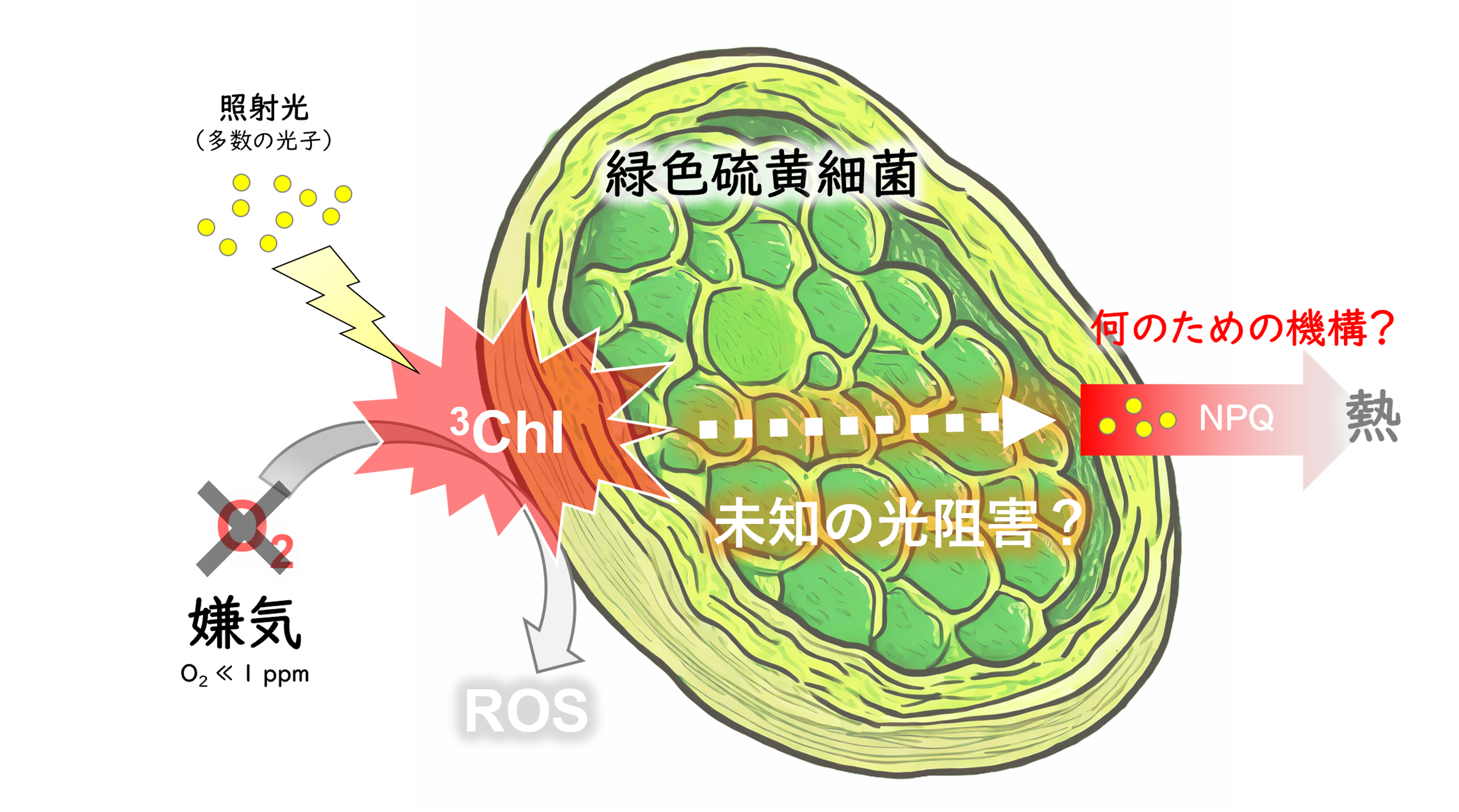
研究キーワード:緑色硫黄細菌,嫌気,カロテノイド,非光化学的消光(NPQ),励起エネルギー移動,原子分解能構造解析,超高速分光
B01: 光合成指数からみる電子伝達反応制御の多様性と進化


概要
光合成は、光エネルギーと水から化学エネルギーと還元力を生み出す「明反応」と、それらを使って二酸化炭素から有機物を作る「暗反応」の共同作業によって成り立つ、というのが教科書に載る光合成の一般概要ですが、厳密には少し違います。とりわけ藻類などでは明反応で生じた還元力の多くが、暗反応とは異なる代謝で消費されており、この割合を示す光合成指数と呼ばれるパラメータは、その生物が「何を目的に光合成を行うのか」に強く関係します。私たちは、多種多様な環境に生息する生物の光合成指数評価を通して、光合成ユビキティの解明に貢献します。
研究キーワード:光合成指数、電子伝達系、カルビン回路、多様性、進化
B01: 海洋の光合成ユビキティを支える珪藻の光捕集システムの機能と進化
代表者

伊福 健太郎(京都大学 大学院農学研究科 教授)![]()
研究協力者

秋本 誠志(神戸大学 大学院理学研究科 准教授)![]()
概要
珪藻は、海洋における光合成の約4分の1を担うともされる重要な微細藻類です。しかし珪藻の光合成システムは緑色植物とは大きく異なり、未解明な点が多く残されています。そこで本研究では、海洋珪藻ツノケイソウ(Chaetoceros gracilis)の光環境応答・適応の分子機構を解析します。様々な光環境条件における網羅的な遺伝子発現情報と立体構造情報を組み合わせ、光環境応答・適応に重要と思われる集光性色素タンパク質(LHC)を絞り込み、それらの遺伝子欠損変異株あるいは過剰発現株を用いて分子機能を解明します。さらに、それら重要LHCの、紅藻、および紅藻を起源とする二次共生藻類における機能保存性を、分子系統学、構造生物学的知見から検討し、海洋における光合成ユビキティを支える紅色進化系統藻類の光捕集系の作動原理と進化を理解することを目指します。
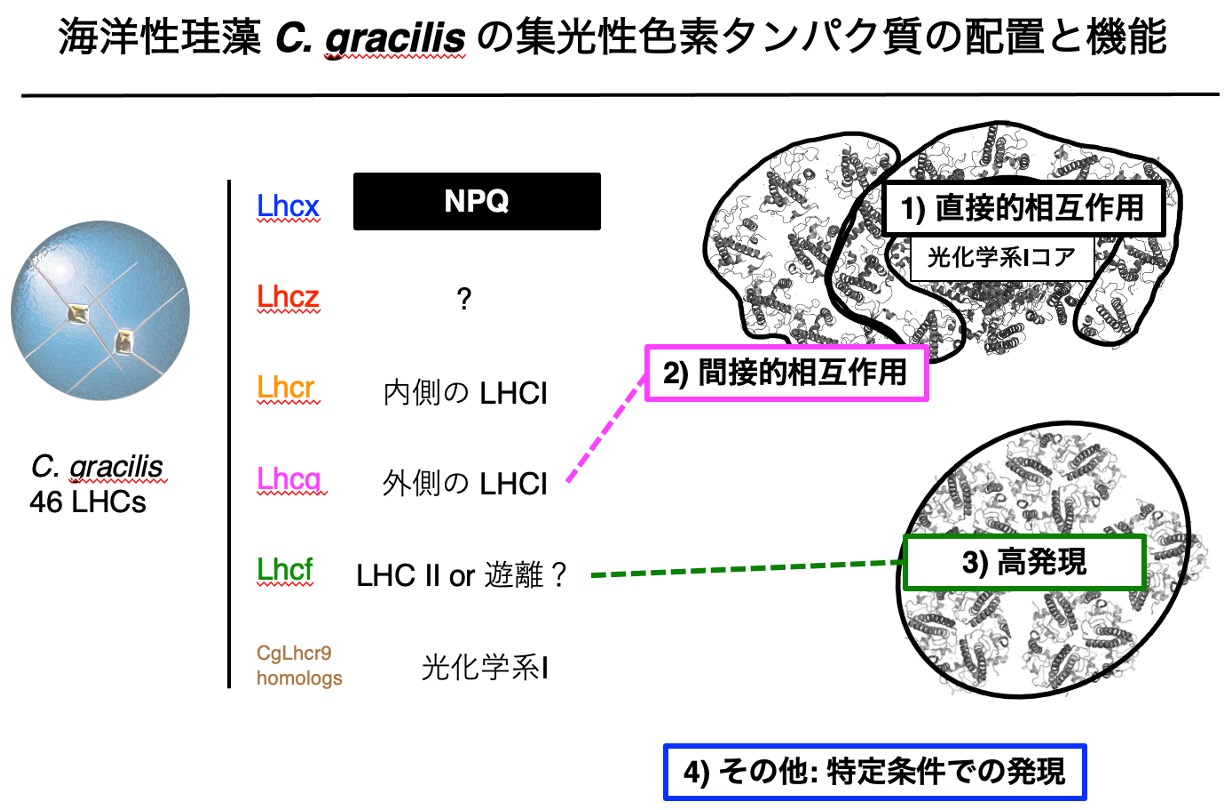
研究キーワード:紅色系統藻類, 2次共生, 集光性色素タンパク質, 環境適応, 分子進化
B01: 近赤外光で駆動する光合成超分子構造の解明
代表者

川上 恵典(理化学研究所 放射光科学研究センター 研究員)![]()
研究協力者

米倉 功治(理化学研究所 放射光科学研究センター グループディレクター)![]()
研究協力者

眞木(米倉)さおり(理化学研究所 放射光科学研究センター 研究員)
研究協力者

内藤 久志(理化学研究所 放射光科学研究センター 研究員)
研究協力者

菓子野 康浩(兵庫県立大学 理学研究科 准教授)
研究協力者

小澄 大輔(熊本大学 産業ナノマテリアル研究所 准教授)
概要
酸素発生型光合成の光化学反応は、膜蛋白質複合体が埋め込まれた「チラコイド膜」上で行われる。生物種ごとで異なる集光性アンテナ複合体がその周りに存在し、太陽光エネルギーを効率的に吸収している。本研究では、クロロフィル (Chl) dを主要色素とするAcaryochloris marina (A. marina)が如何にして近赤外光を利用して光合成を行うのか、その全容を明らかにするため、クライオ電子顕微鏡(Cryo-EM)によるA. marina由来「水溶性アンテナ複合体-光化学系膜蛋白質」の立体構造解析を行う。さらに、クライオ電子線トモグラフィー(Cryo-ET)によってA. marina細胞内のチラコイド膜構造を三次元的に可視化し、in vivoでのアンテナ複合体と光化学系蛋白質との相互作用の理解を目指す。
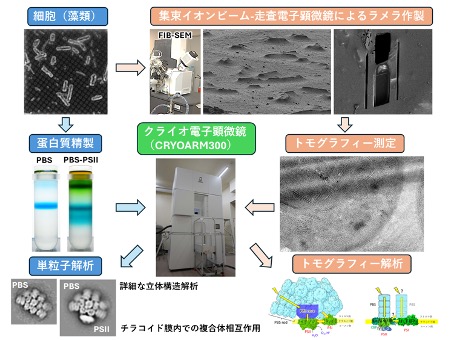
研究キーワード:シアノバクテリア、タンパク質精製、クライオ電子顕微鏡、単粒子解析、クライオ電子トモグラフィー、構造解析
B01: 光合成の多様性を支える電子受容体NADP供給系の変容
代表者
 川合 真紀(埼玉大学 理工学研究科 准教授)
川合 真紀(埼玉大学 理工学研究科 准教授)![]()
研究協力者

宮城 敦子(山形大学 農学部 准教授)
研究協力者

橋田 慎之介(電力中央研究所 サステナブルシステム研究本部 上席研究員)
概要
NAD(P)(H)は、全ての生物が様々な細胞内酸化還元反応に用いる電子伝達物質です。中でもNADP+は光合成リニア電子伝達における最終電子受容体として機能することから、光合成生物はこれを光合成マシーナリーに効率よく供給する仕組みを獲得していると考えられます。本研究では、様々な環境に適応した光合成生物における光合成リニア電子伝達鎖への電子受容体供給システムの多様性とその分子メカニズムを葉緑体内のNAD(P)(H)代謝の観点から解明します。
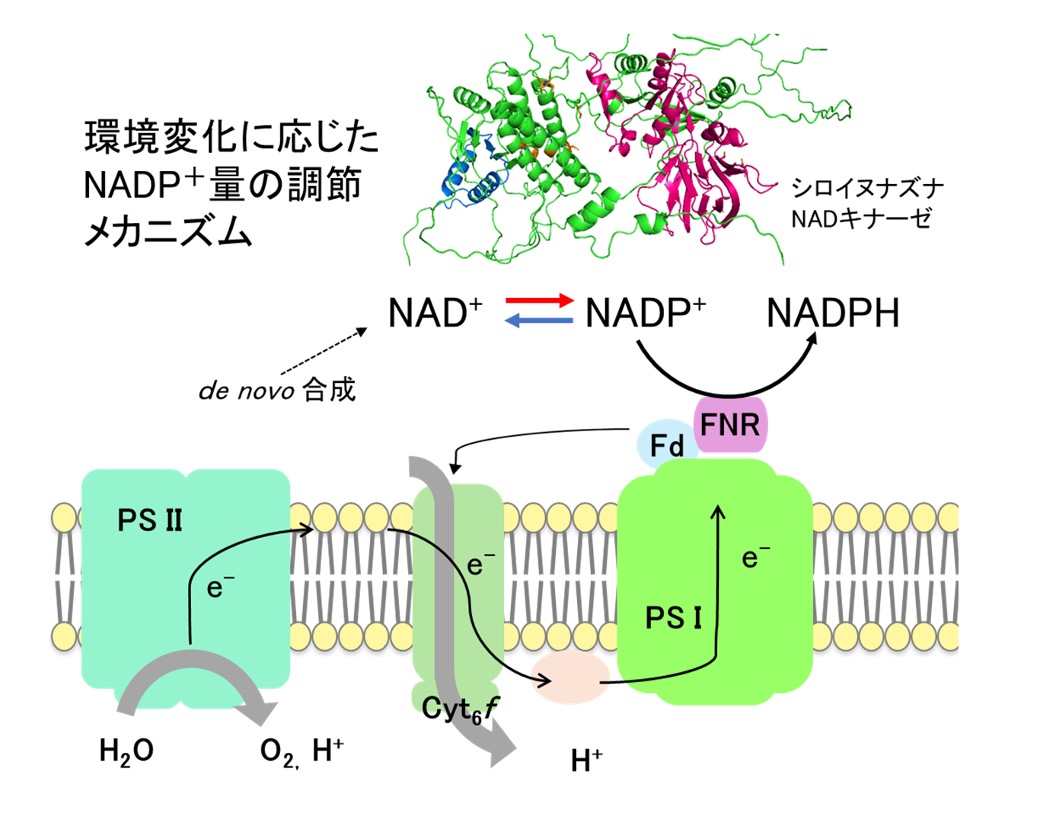
研究キーワード:葉緑体、光合成、NAD(P)(H)、NADキナーゼ、NADPホスファターゼ、酵素活性調整
B01: 海洋性大型緑藻の集光蛋白質における色素の結合部位による光応答制御機構の解明
代表者

藤井 律子(大阪公立大学人工光合成研究センター 准教授)![]()
研究協力者

オッフィーヨアンダラ アヤル(大阪公立大学人工光合成研究センター 研究員)
研究協力者

関 荘一郎(大阪大学 蛋白質研究所 研究員)
概要
高等植物や緑藻において集光蛋白質は、太陽光を集める集光と過剰なエネルギーを安全に逃す消光という二つの役割を備え、様々な自然光に対して反応中心をうまく駆動する励起エネルギーを供給しています。高等植物の集光蛋白質LHCIIについては、集光/消光の機能が色素のタンパク質中での周辺環境により絶妙に制御されていることがわかってきましたが、特定の色素を特定の結合サイトに選択的に結合する仕組みについてはほとんどわかっていません。そこで本研究では、高等植物のLHCIIとほぼ同じ蛋白質を持つが色素が異なっているLHCIIを持つシフォナス緑藻をターゲットとし、クライオ電顕法により構造生物学的に色素の結合サイトを解明し、同時に分光学的な応答に関する知見を蓄積して、色素の結合部位を制御する要因を明らかにすることを目指します。
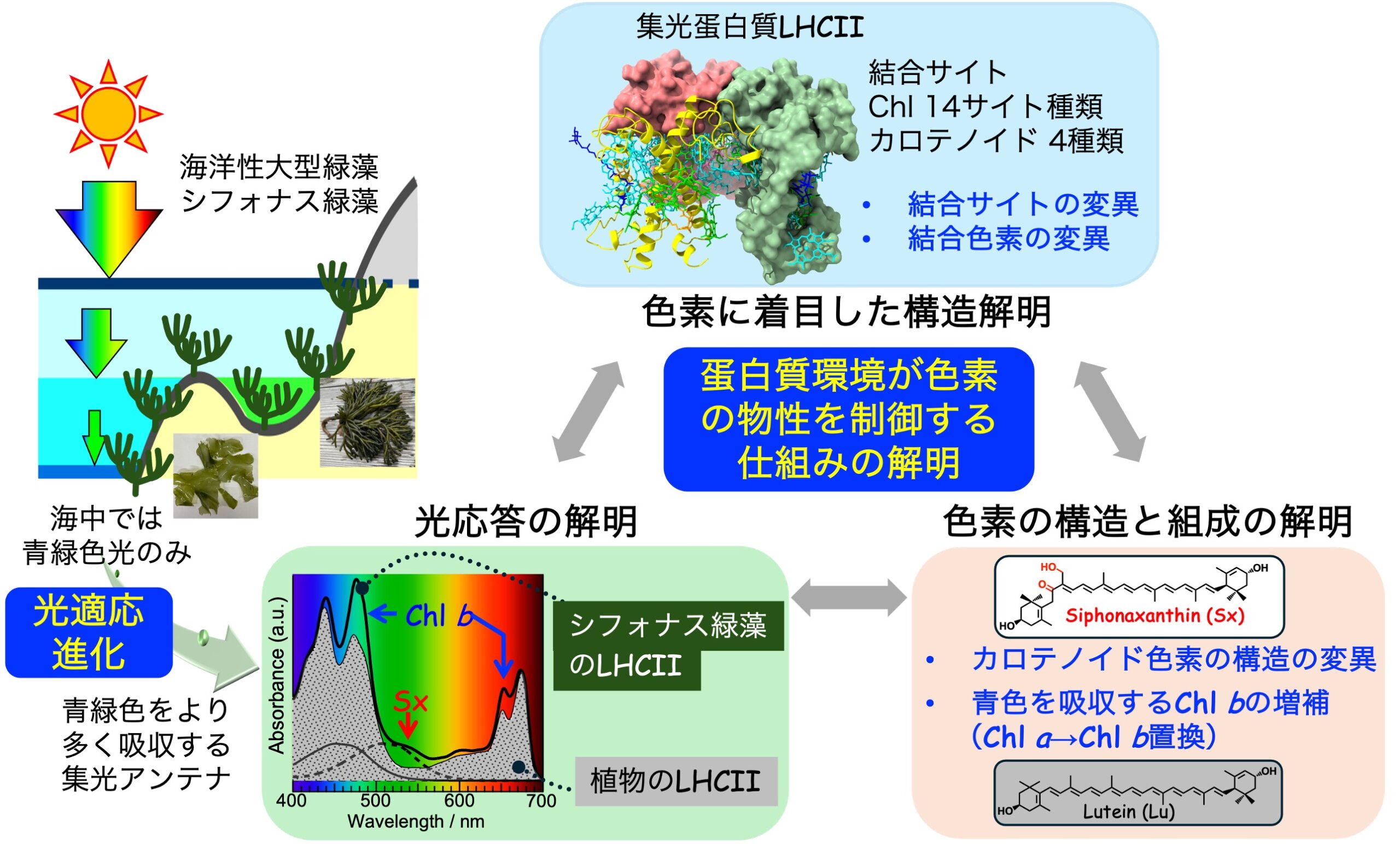
研究キーワード:緑藻、LHCII、シフォナキサンチン(カロテノイド)、色素分析、分光分析、クライオ電顕
B01: 維管束鞘細胞葉緑体グラナフリーチラコイド膜上の光化学系超分子複合体形成の分子機構
代表者
 宗景 ゆり(関西学院大学 生命環境学部生物科学科 教授)
宗景 ゆり(関西学院大学 生命環境学部生物科学科 教授)![]()
研究協力者
 大井 崇生(高知工科大学 理工学群 准教授)
大井 崇生(高知工科大学 理工学群 准教授)
概要
C4植物には維管束鞘細胞葉緑体において光化学系IIの活性抑制と共にグラナ構造が消失したチラコイド膜構造を有するタイプがあります。この構造は、光化学系Iの集光効率を高めることで、環状電子伝達によるATP合成を促進することが予想されます。本研究では、グラナ構造が消失したチラコイド膜上に形成される光化学系超複合体において、光エネルギーの移動がどのように起こるのか、また、この超複合体とチラコイド構造の形成にどのような分子機構が関与しているのかを明らかにすることで、C4植物の進化の過程で生じた光合成最適化システムを解明することを目指します。
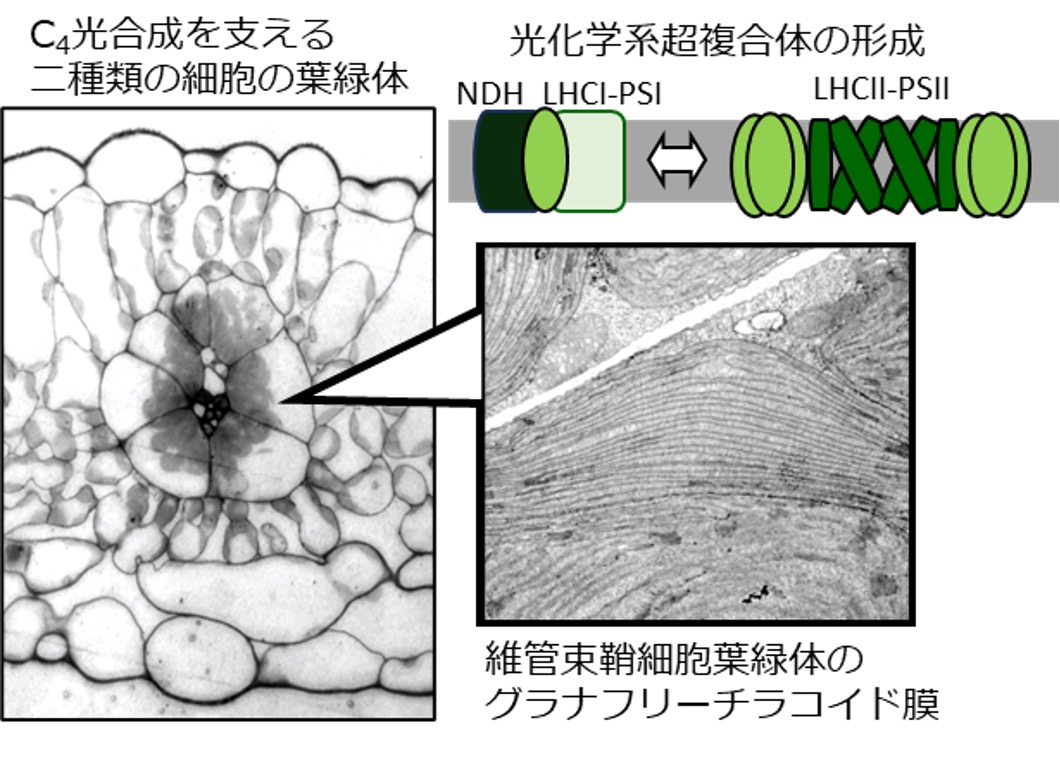
研究キーワード:C4植物、進化、チラコイド膜構造、循環型電子伝達、光化学系超複合体
B01: 過酷な生育環境を生き抜く微細藻類が持つユニークな光酸化ストレス防御機構の解明
代表者

川﨑 信治(東京農業大学 生命科学部分子微生物学科 教授)![]()
研究協力者

庄村 康人(茨城大学 大学院理工学研究科 准教授)
研究協力者
豊島 拓樹(東京農業大学 生命科学部分子微生物学科 助教)
概要
一般植物は、強光照射下で、かつ乾燥や塩ストレスなどが混在する過酷な環境下では、光酸化ストレスを発生し枯死に至ります。我々は上記の環境を生き抜く光合成微生物を探索し、単離株が持つユニークなストレス防御機構の研究を行ってきました。その結果、単離株は高い光酸化ストレス耐性を示し、報告例の無いユニークな細胞応答を保持していました。本申請では、ユニークな生理応答を保持する単離株の分子機構を多角的な専門分野からアプローチし、未知の光酸化ストレス防御機構の解明を目指します。
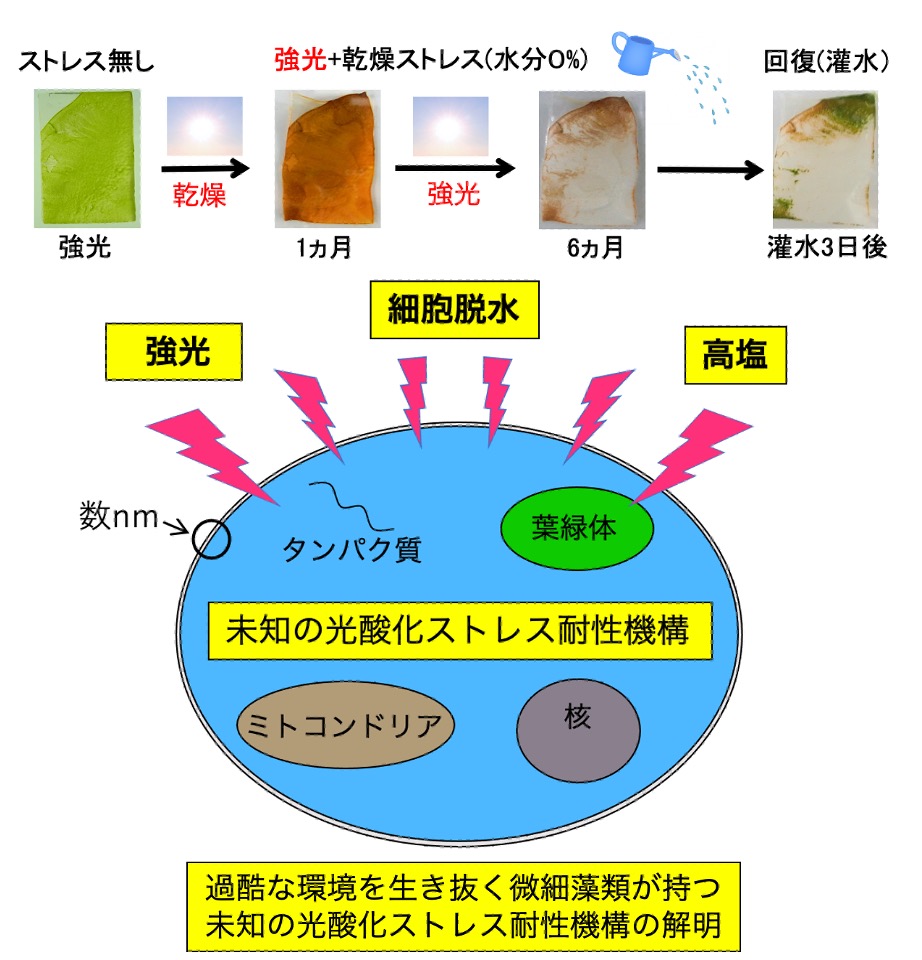
研究キーワード:微細藻類、光酸化ストレス、カロテノイド、アスタキサンチン、活性酸素
B01: 金属配位酵素の分配制御による光捕集系構築の分子機構の解明
代表者

増田 建(東京大学大学院総合文化研究科 教授)![]()
研究協力者

三宅 敬太(東京大学大学院総合文化研究科 助教)![]()
概要
本研究では、金属配位酵素の分配制御による光捕集系構築の分子機構の解明を目的とする。光合成ユビキティにおいて、光エネルギー捕集系の構築は必須である。光合成生物の光捕集系は生育する光環境に応じて多様であるが、陸上植物、緑藻、原核緑藻が有する、光捕集クロロフィルタンパク質(LHC)系と、原核緑藻を除くシアノバクテリアや紅藻に代表される、ビリン色素を主とするフィコビリソーム(PBS)系に大別出来る。これら光捕集系の構築には、大量の光合成色素の供給が必須であり、これはポルフィリンから金属配位酵素への分岐段階での分配制御に依存する。本研究ではLHC系およびPBS系のシアノバクテリアを実験材料として、光捕集系への光合成色素供給の観点から、金属配位段階の制御について、1.LHC系とPBS系の光合成色素合成関連遺伝子のバイオインフォマティクスによる比較ゲノム解析。2.遺伝子機能解析、3.金属配位酵素を含むクロロフィルおよびヘム・ビリン合成系のin vitroフロー解析により、その分子機構の解明に取り組む。
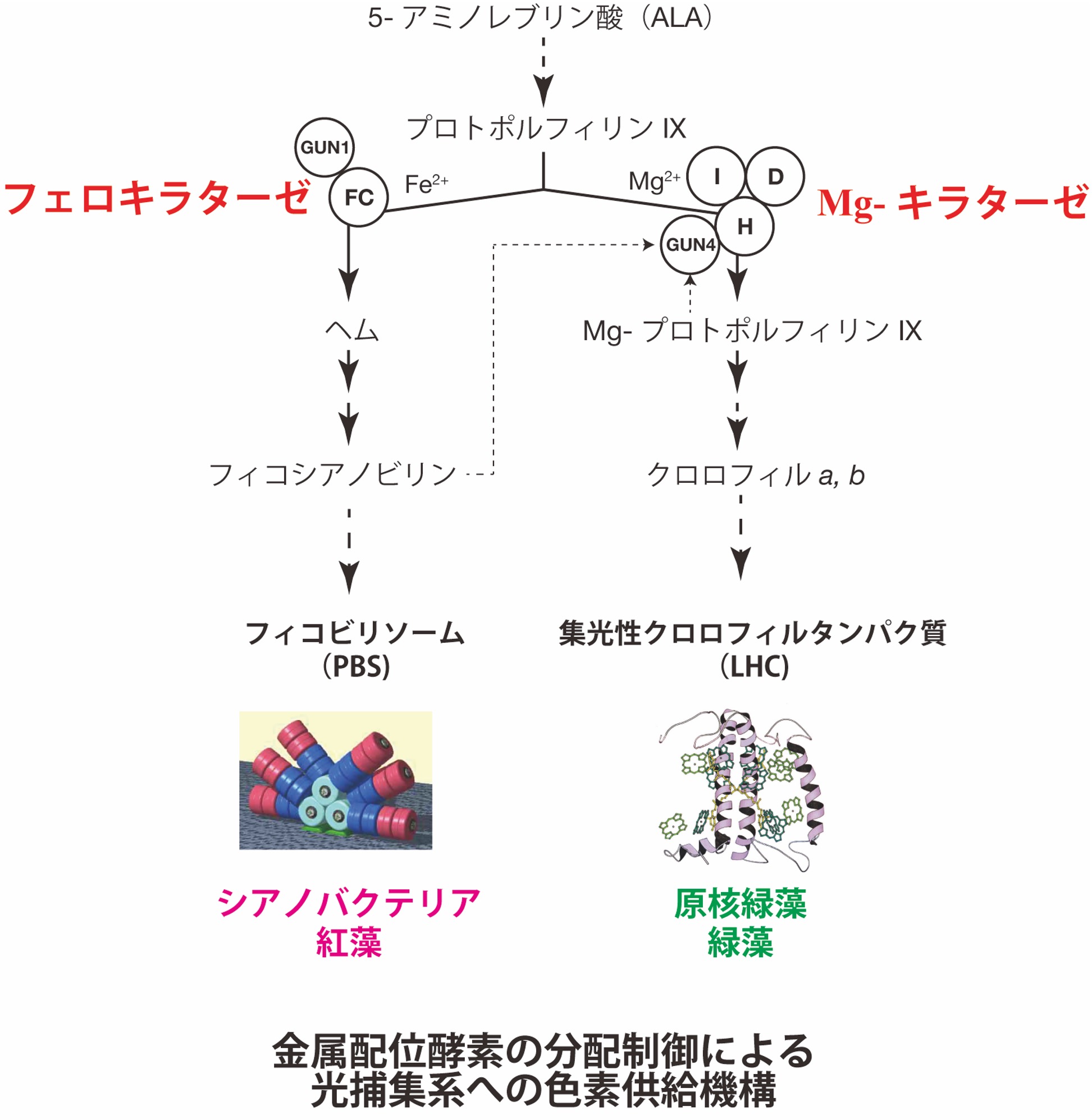
研究キーワード:金属配位酵素,クロロフィル,ビリン,光捕集系,テトラピロール、in vitro 再構成
B01: 紅色硫黄光合成細菌におけるカルシウムを利用した環境適応戦略の解明
代表者
 木村 行宏(神戸大学大学院農学研究科 准教授)
木村 行宏(神戸大学大学院農学研究科 准教授)![]()
研究協力者
大友 征宇(茨城大学理学部 教授)
研究協力者
谷 一寿(筑波大学計算科学研究センター 教授)
概要
紅色光合成細菌は近赤外光を光電変換する光捕集・電荷分離システム(光捕集1反応中心複合体:LH1-RC)を備えており、LH1からRCへのuphill型エネル ギー移動により近赤外光をエネルギー源とした光—物質変換を営んでいる。本申請者らは、Chromatiaceae科の紅色硫黄細菌がカルシウムを利用して温度や光に対する環境適応能力を獲得してきたという仮説を提唱している。本研究では、 近年、日本で発見された好熱性の紅色硫黄細菌Cardichlormatium japonicum由来LH1-RCの構造と機能におけるカルシウムの役割を網羅的に解析し、他のChromatiaceaeとの比較研究を行うことにより、Chromatiaceae科におけるカルシウムを利用した環境適応および多様化の生存戦略を明らかにする。
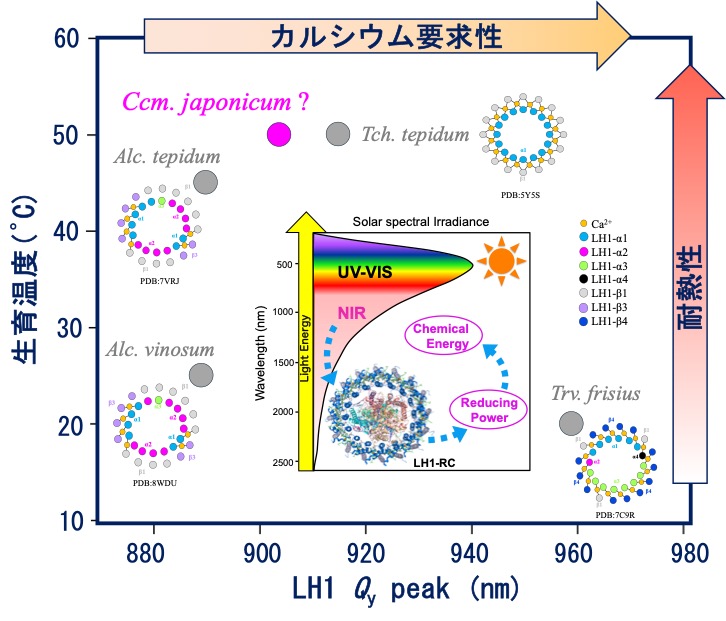
研究キーワード:光合成細菌、光捕集1反応中心複合体、カルシウム、近赤外光、分光分析、熱量分析、構造解析
B01: 赤潮形成藻シャットネラ属の光合成環境適応機構
代表者

西山 佳孝(埼玉大学大学院理工学研究科 准教授)![]()
研究協力者

神保 晴彦(埼玉大学大学院理工学研究科 助教)
研究協力者

湯浅 光貴(水産研究・教育機構水産技術研究所 研究員)
概要
有害な⾚潮を形成するラフィド藻シャットネラ属は、強光・⾼温に対する耐性が強く、夏場に海⾯直下でブルームを形成して養殖⿂を斃死させる。シャットネラ属の光合成は強光・⾼温に対する耐性が極めて高く、特殊な光合成制御機構を有していることが推測される。本研究では、シャットネラ属の光合成の強光・⾼温耐性のメカニズムを解明することを⽬的とする。
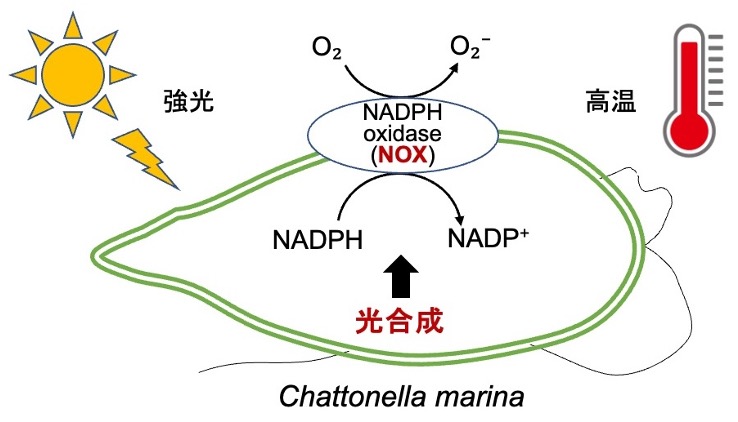
研究キーワード:赤潮、シャットネラ、強光耐性、高温耐性、光合成測定、活性酸素測定
B01: 葉の形態と葉色の多様性に伴う葉温調節と光合成反応制御
代表者

滝澤 謙二(アストロバイオロジーセンター 特任准教授)![]()
概要
光合成生物は地球上の多様な環境下で光合成を行うために、様々な分子機構を進化させてきました。陸上植物では自然環境と光合成反応の場である葉緑体とを隔てて重層的な葉肉組織が存在します。本公募班では多様な葉の形態と色(色素)により形成される葉内環境(特に葉内温度)が外部の過酷な自然環境を和らげる緩衝作用を明らかにするとともに、光合成反応の葉内環境への適応メカニズムの解明を目指します。
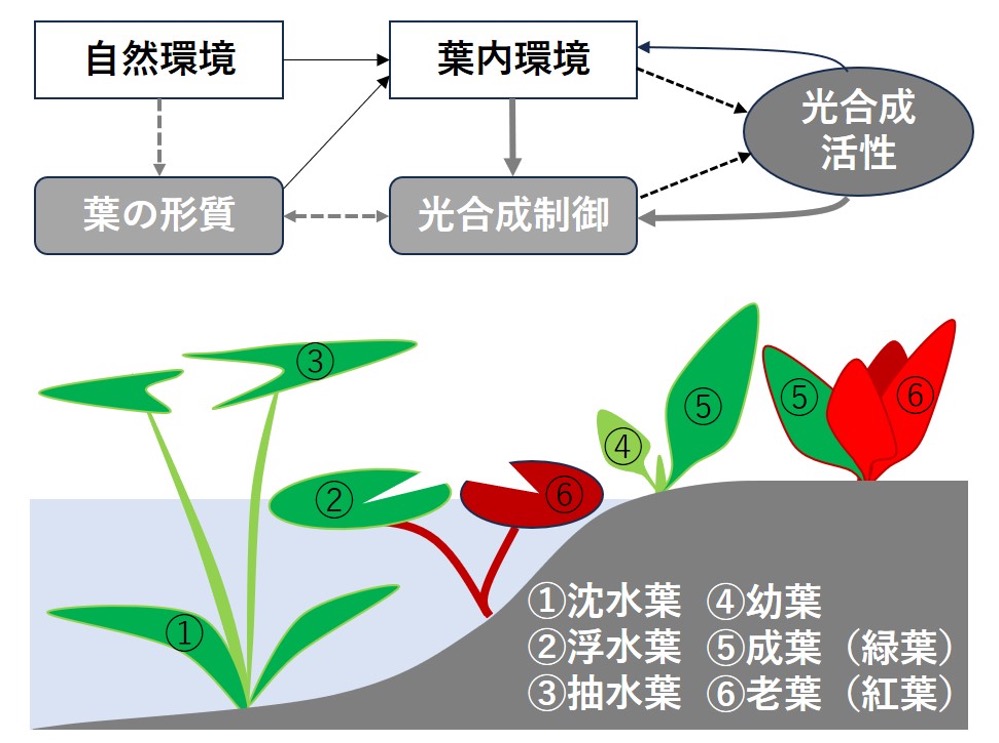
研究キーワード:葉肉組織、フラボノイド、温度調節、陸上化
B01: 葉緑体ATP合成酵素の機能的多様性の構造・進化的理由
代表者

上妻 馨梨(京都大学 大学院農学研究科 助教) ![]()
研究協力者
伊福 健太郎(京都大学 大学院農学研究科 教授)概要
葉緑体ATP合成酵素のレドックス制御は非常にユニークです。酵素の回転軸であるγsubunit(ATPC)が持つ1対のシステインは地上部(ATPC1)では酵素のon/offの機能を制御しますが、地下部(ATPC2)ではその機能を持たず常に活性型(on)を保持します。地下部で不活性型(off)にならない理由は構造が原因であると考えられますが、何故、地上部と地下部で制御が異なるのか?光のない地下部での機能は何か?構造の違いと局在は進化の過程でどのように分配されたのか?γ subunitを取り込む他のサブユニットも制御されているのか?個体内における葉緑体ATP合成酵素のレドックス制御の多様性とそれを獲得した構造的理由、さらには進化的理由を解明します。
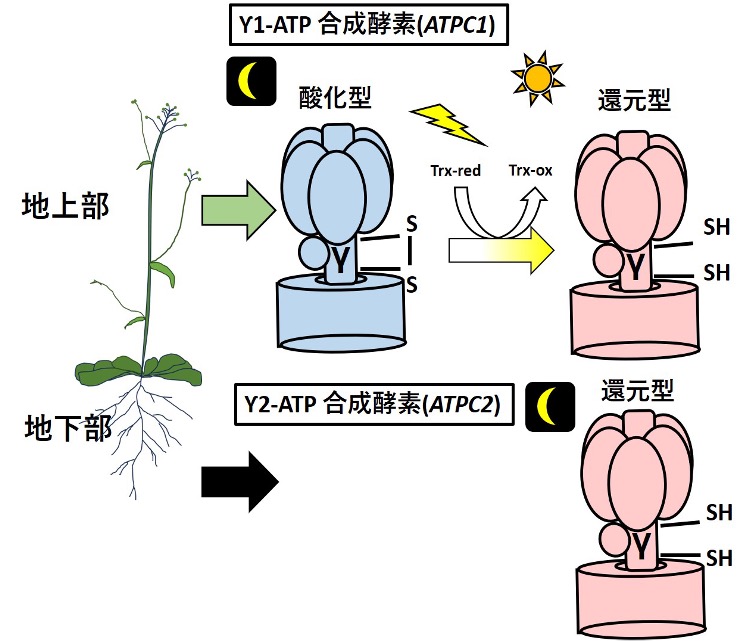
研究キーワード:植物、チラコイド膜電位、葉緑体ATP合成酵素、レドックス制御、分光反射
B02: 高い波長分解能を持つ超高速分光測定による光合成超分子複合体の詳細機能解析
代表者
秋本 誠志(神戸大学 大学院理学研究科 准教授) ![]()
概要
近年、光化学系I、光化学系II、アンテナ色素タンパク質、および、これらから形成される超複合体の構造が次々に明らかにされつつあります。さらに、色素–色素間相互作用や色素–タンパク質間相互作用を評価しタンパク質中の各色素のエネルギーが求められれば、エネルギー移動経路や消光のメカニズムを理論的に予測できる状況になりつつあります。このような学術的背景のもと、本研究では、高いエネルギー分解能を持った超高速分光法により、光合成超複合体の機能を解析するための基盤を構築します。時間–波長の高分解能測定データを得て、エネルギー移動・消光過程を議論していきます。
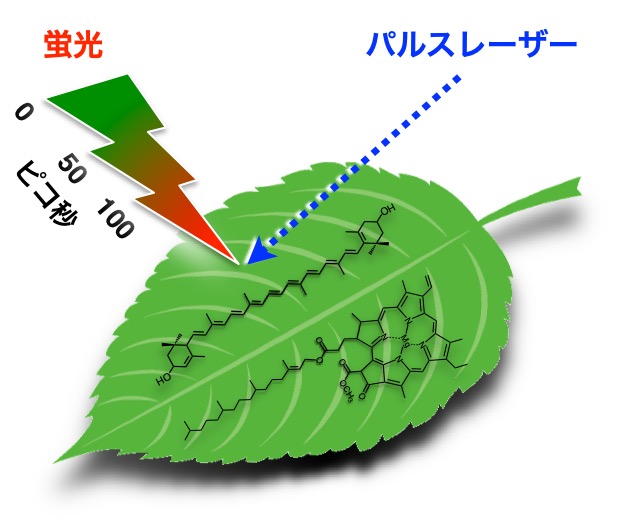
研究キーワード:時間分解分光法、蛍光、エネルギー移動、消光
B02: 単粒子分光解析による過渡的光合成超複合体のエネルギー移動過程の解明
代表者

柴田 穣(東北大学大学院 理学研究科 准教授) ![]()
研究協力者
 谷口 凛(東北大学大学院 理学研究科 博士後期課程)
谷口 凛(東北大学大学院 理学研究科 博士後期課程)概要
巨大な光合成超分子複合体の構造解析を可能とすることで,クライオ電子顕微鏡の単粒子構造解析の発展が光合成研究に革新をもたらしている。少ない標品で解析が可能なことから,近い将来には過渡的にしか蓄積しない希少分子種の構造も解明されると予想される。しかし,光合成機能の中核は光吸収に続くエネルギー移動や電荷分離などの一連の光反応であり,分光解析技術は変わらず重要である。一方で,分光解析には純度の高い均一な標品が比較的多く必要とされるため,構造情報は得られるが分光情報が得られない,という課題が今後生じる可能性がある。希少で多少の不均一性もある標品でも,高度な分光解析が可能となる手法が必要とされている。本研究では,申請者がこれまで推進してきた単一粒子分光法の高度化をさらに発展させ,希少分子種の励起スペクトル測定およびピコ秒時間分解蛍光スペクトル測定を可能とする装置開発を行う。この実現により,光捕集の制御過程や超複合体の組み立て(Assembly)過程に生じる過渡的分子種のエネルギー移動や光防御機構としての消光過程を明らかにする。
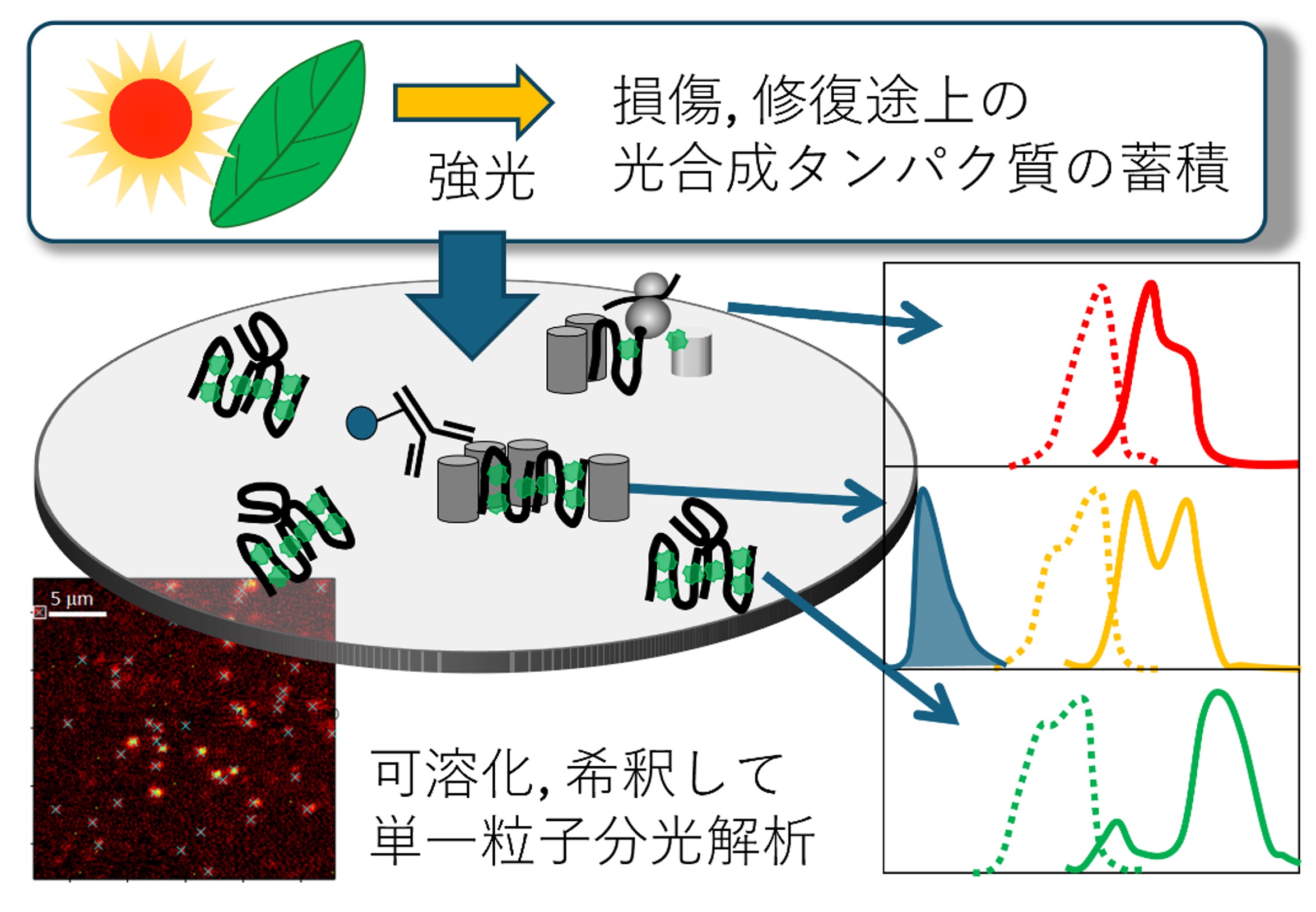
研究キーワード:単一粒子分光、蛍光寿命解析、極低温共焦点顕微鏡
B02: スピン拡散法による光合成酸素発生系の磁気構造解析
代表者

三野 広幸(名古屋大学 理学研究科 准教授) ![]()
概要
光合成酸素発生反応は光化学系Ⅱタンパク質のマンガンクラスターで行われる。反応中間状態の分子構造はX線結晶構造解析により構造が解明されつつあるが、分子構造の解明だけでは酸素発生メカニズムは解明できない。本研究はマンガンクラスターの高酸化中間状態S2、S3の電子状態を電子スピン共鳴(ESR)により解析し、酸素発生過程を解明することを目的としている。また、本研究ではこれまでESR信号として観測できなかった高スピン状態の酸素発生系の全中間状態の磁気的構造をスペクトル拡散法によって実験的に測定し、マンガンクラスターの構造及び分子軌道を解析する。
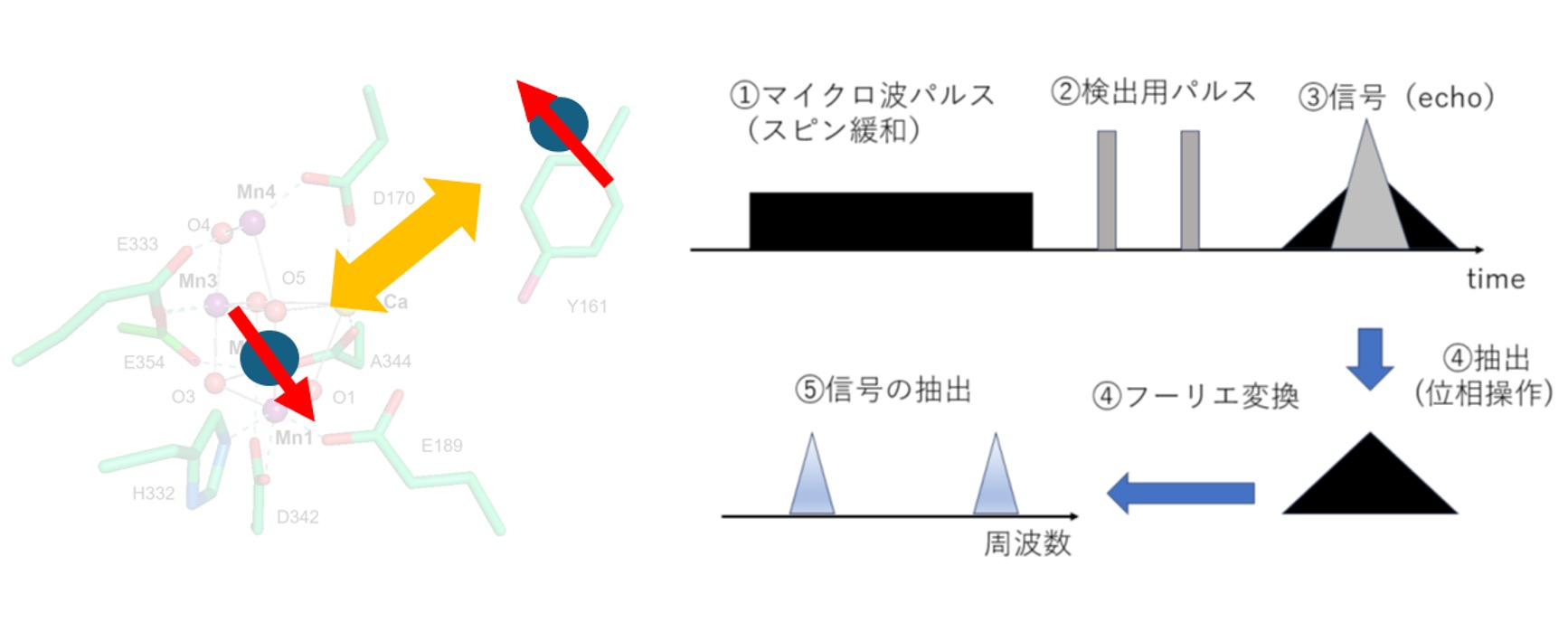
研究キーワード:酸素発生系、マンガンクラスター、電子スピン共鳴、量子化学計算
B02: 紅色細菌のクロマトフォア膜の in situ 構造生物学
代表者
 山形 敦史(理化学研究所 上級研究員)
山形 敦史(理化学研究所 上級研究員) ![]()
研究協力者
 ローラ・ブラカン(理化学研究所 研究員)
ローラ・ブラカン(理化学研究所 研究員)概要
ロドバクター類は赤褐色を示す光合成細菌(紅色細菌)の一つです。ロドバクターの光合成は細胞内のクロマトフォア小胞で行われます。クロマトフォア小胞の膜では光反応中心(RC)・光捕集タンパク質I(LH1)複合体及び光捕集タンパク質II(LHII)が高密度に配置され、光エネルギーの捕集や受け渡しが最適化されていると考えられます。本研究では、クライオ電子顕微鏡を用いてRC-LH1やLHIIといった光合成タンパク質群を周辺環境を含んだまま可視化する in situ 構造解析を行います。粗精製したクロマトフォア小胞膜の可視化に加えて、ワッフル法と呼ばれる新しい高圧凍結法とクライオ収束イオンビーム加工を組み合わせて菌体内のクロマトフォア小胞を可視化することに挑戦します。
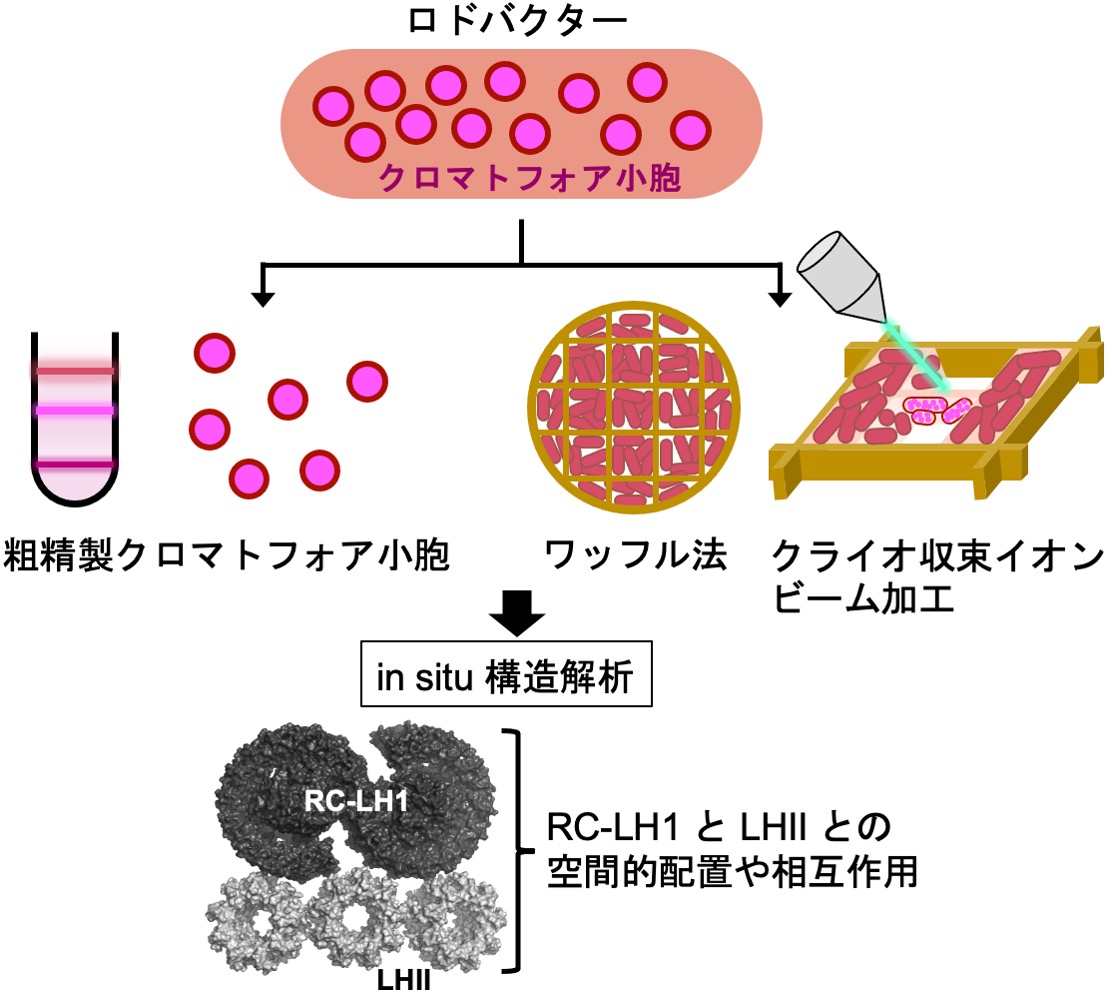
研究キーワード:ロドバクター、光反応中心・光捕集タンパク質複合体、”その場”(in situ)構造解析、クライオ電子線トモグラフィー、クライオ電子顕微鏡、高圧凍結、クライオ収束イオンビーム加工
B02: シアノバクテリオクロムの光変換の構造基盤
代表者
![]() 三島 正規(東京薬科大学 薬学部 教授)
三島 正規(東京薬科大学 薬学部 教授) ![]()
研究協力者
 青山 洋史(東京薬科大学 薬学部 准教授)
青山 洋史(東京薬科大学 薬学部 准教授)研究協力者
 武田 光広(東京薬科大学 薬学部 助教)
武田 光広(東京薬科大学 薬学部 助教)研究協力者
 永江 峰幸(東京薬科大学 薬学部 助教)
永江 峰幸(東京薬科大学 薬学部 助教)概要
シアノバクテリアがもつフィトクロム様タンパク質(シアノバクテリオクロム)は,発色団フィコシアノビリン(PCB)の結合によって紫外光から赤外光の幅広い波長領域に応答する光センサータンパク質である.光吸収に伴うPCB分子内の二重結合の異性化は,GAF・PASドメインからなるリンカー領域を経て最終的にヒスチジンキナーゼ(HK)ドメインへとアロステリックに伝達され,キナーゼ活性が制御される.本研究では,光捕集アンテナであるフィコビリソームの構成を調節する緑/赤色光吸収型のシアノバクテリオクロムであるRcaEタンパク質の光吸収機構(多様な極大吸収波長は如何にもたらされるか)と,アロステリックな構造変化伝達機構を,X線結晶構造解析およびNMR法を主たる手法として,原子分解能で解明する.全長の解析ではクライオ電子顕微鏡単粒子解析,光センサー部であるGAFドメインについては中性子線結晶構造解析へと研究を展開する.
![]()
研究キーワード:シアノバクテリオクロム、NMR、X線結晶構造解析、有機合成